How
we got here, and where we are going. Ever since the writings of Darwin and Huxley,
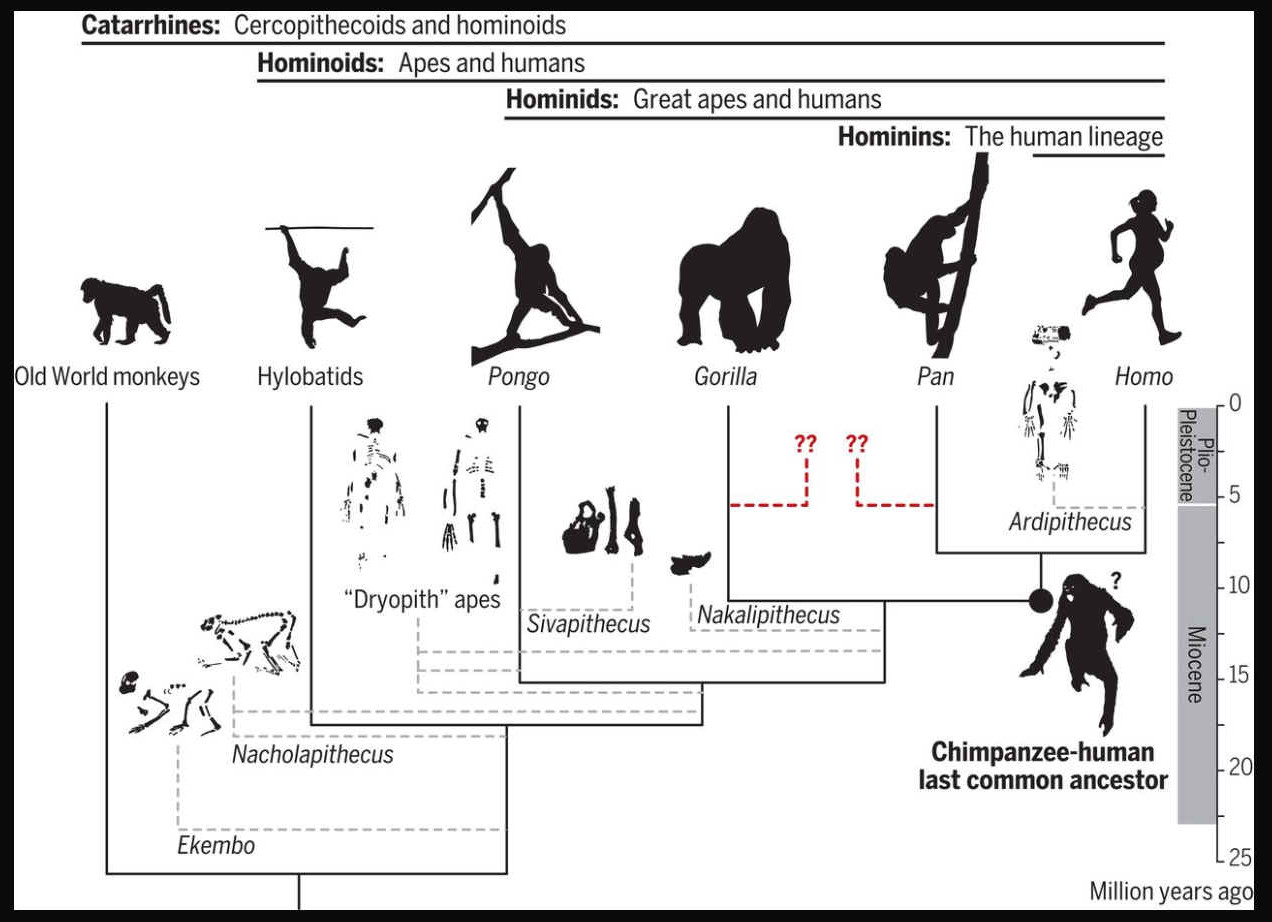
humans’ place in nature relative to apes (nonhuman hominoids) and the geographic origins of the human lineage (hominins) have been heavily debated. Humans diverged from apes [specifically, the chimpanzee lineage (Pan)] at some point between ~9.3 million and ~6.5 million years ago (Ma), and habitual bipedalism evolved early in hominins (accompanied by enhanced manipulation and, later on, cognition). To understand the selective pressures surrounding hominin origins, it is necessary to reconstruct the morphology, behavior, and environment of the Pan-Homo last common ancestor (LCA). “Top-down” approaches have relied on living apes (especially chimpanzees) to reconstruct hominin origins. However, “bottom-up” perspectives from the fossil record suggest that modern hominoids represent a decimated and biased sample of a larger ancient radiation and present alternative possibilities for the morphology and geography of the Pan-Homo LCA. Reconciling these two views remains at the core of the human origins problem.
Homo heidelbergensis (also H. sapiens heidelbergensis) is an extinct species or subspecies of archaic human which existed during the Middle Pleistocene. It was subsumed as a subspecies of H. erectus in 1950 as H. e. heidelbergensis, but towards the end of the century, it was more widely classified as its own species. It is debated whether or not to constrain H. heidelbergensis to only Europe or to also include African and Asian specimens, and this is further confounded by the type specimen (Mauer 1) being a jawbone, because jawbones feature few diagnostic traits and are generally missing among Middle Pleistocene specimens. Thus, it is debated if some of these specimens could be split off into their own species or a subspecies of H. erectus. Because the classification is so disputed, the Middle Pleistocene is often called the "muddle in the middle."
H. heidelbergensis is regarded as a chronospecies, evolving from an African form of H. erectus (sometimes called H. ergaster). By convention, H. heidelbergensis is placed as the most recent common ancestor between modern humans (H. sapiens or H. s. sapiens) and Neanderthals (H. neanderthalensis or H. s. neanderthalensis). Many specimens assigned to H. heidelbergensis likely existed well after the modern human/Neanderthal split. In the Middle Pleistocene, brain size averaged about 1,200 cubic centimetres (cc), comparable to modern humans. Height in the Middle Pleistocene can only be estimated based upon remains from 3 localities: Sima de los Huesos, Spain, 169.5 cm (5 ft 7 in) for males and 157.7 cm (5 ft 2 in) for females; 165 cm (5 ft 5 in) for a female from Jinniushan, China; and 181.2 cm (5 ft 11 in) for a specimen from Kabwe, Zambia. Like Neanderthals, they had wide chests and were robust overall.
The Middle Pleistocene of Africa and Europe features the advent of Late Acheulian technology, diverging from that of earlier and contemporary H. erectus, and probably issuing from increasing intelligence. Fire likely became an integral part of daily life after 400,000 years ago, and this roughly coincides with more permanent and widespread occupation of Europe (above 45°N), and the appearance of hafting technology to create spears. H. heidelbergensis may have been able to carry out coordinated hunting strategies, and consequently they seem to have had a higher dependence on meat.

EVOLUTION
H. heidelbergensis is thought to have descended from African H. erectus — sometimes classified as Homo ergaster — during the first early expansions of hominins out of Africa beginning roughly 2 million years ago. Those that dispersed across Europe and stayed in Africa evolved into H. heidelbergensis or speciated into H. heidelbergensis in Europe and "H. rhodesiensis" in Africa, and those that dispersed across East Asia evolved into H. erectus s. s. The exact derivation from an ancestor species is obfuscated by a long gap in the human fossil record near the end of the Early Pleistocene. In 2016, Antonio Profico and colleagues suggested that 875,000-year-old skull materials from the Gombore II site of the Melka Kunture Formation, Ethiopia, represent a transitional morph between H. ergaster and H. heidelbergensis, and thus postulated that H. heidelbergensis originated in Africa instead of Europe.
According to genetic analysis, the LCA of modern humans and Neanderthal split into a modern human line, and a Neanderthal/Denisovan line, and the latter later split into Neanderthal and Denisovans. According to nuclear DNA analysis, the 430,000-year-old SH humans are more closely related to Neanderthals than Denisovans (and that the Neanderthal/Denisovan, and thus the modern human/Neanderthal split, had already occurred), suggesting the modern human/Neanderthal LCA had existed long before many European specimens typically assigned to H. heidelbergensis did, such as the Arago and Petralona materials.
In 1997, Spanish archaeologist José María Bermúdez de Castro, Arsuaga, and colleagues described the roughly million-year-old H. antecessor from Gran Dolina, Sierra de Atapuerca, and suggested supplanting this species in the place of H. heidelbergensis for the LCA between modern humans and Neanderthals, with H. heidelbergensis descending from it and being a strictly European species ancestral to only Neanderthals. They later recanted. In 2020, Dutch molecular palaeoanthropologist Frido Welker and colleagues analysed ancient proteins collected from an H. antecessor tooth found that it was a member of a sister lineage to the LCA rather than being the LCA itself (that is, H. heidelbergensis did not derive from H. antecessor).
Human dispersal beyond 45°N seems to have been quite limited during the Lower Palaeolithic, with evidence of short-lived dispersals northward beginning after a million years ago. Beginning 700,000 years ago, more permanent populations seem to have persisted across the line coinciding with the spread of hand axe technology across Europe, possibly associated with the dispersal of H. heidelbergensis and behavioural shifts to cope with the cold climate. Such occupation becomes much more frequent after 500,000 years ago.
NUTRITION
Middle Pleistocene communities in general seem to have eaten big game at a higher frequency than predecessors, with meat becoming an essential dietary component. Diet could overall be
varied - for example the inhabitants of Terra Amata seem to have been mainly eating deer, but also elephants, boar, ibex, rhino and aurochs. African sites typically commonly yield bovine and horse bones. Though carcasses may have simply been scavenged, some Afro-European sites show specific targeting of a single species, which more likely indicates active hunting; for example: Olorgesailie, Kenya, which has yielded over 50 to 60 individual baboons (Theropithecus oswaldi); and Torralba and Ambrona in Spain which have an abundance of elephant bones (though also rhino and large hoofed mammals). The increase in meat subsistence could indicate the development of group hunting strategies in the Middle Pleistocene. For instance, at Torralba and Ambrona, the animals may have been run into swamplands before being killed, entailing encircling and driving by a large group of hunters in a coordinated and organised attack. Exploitation of aquatic environments is generally quite lacking, despite some sites being in close proximity to the ocean, lakes or rivers.
Plants were probably also frequently consumed, including seasonally available ones, but the extent of their exploitation is unclear as they do not fossilise as well as animal bones. Assuming a diet heavy in lean meat, an individual would have needed a high carbohydrate intake to prevent protein poisoning, such as by eating typically abundant underground storage organs, tree bark, berries, or nuts. The Schöningen site, Germany, has over 200 plants in the vicinity which are either edible raw or when cooked.

HOMO
ERECTUS: artists impression of a female of the species, with flattened
nose and arched brows. Not intellectually well developed, but with many
evolutionary advantages, mainly walking upright and using tools.
ART
Upper Palaeolithic modern humans are well known for having etched
engravings seemingly with symbolic value. As of 2018, only 27 Middle and
Lower Palaeolithic objects have been postulated to have symbolic
etching, out of which some have been refuted as having been caused by
natural or otherwise non-symbolic phenomena (such as the fossilisation
or excavation processes). The Lower Palaeolithic ones are: three
380,000-year-old pebbles from Terra Amata; a 250,000-year-old pebble
from Markkleeberg, Germany; 18 roughly 200,000-year-old pebbles from
Lazaret (near Terra Amata); a roughly 200,000-year-old lithic from
Grotte de l'Observatoire, Monaco; a 370,000-year-old bone from
Bilzingsleben, Germany; and a 200- to 130-thousand-year-old pebble from
Baume Bonne, France.
In the mid-19th century, French archaeologist Jacques Boucher de Crèvecœur
de Perthes began excavation at St. Acheul, Amiens, France, (the area
where the Acheulian was defined), and, in addition to hand axes,
reported perforated sponge fossils (Porosphaera globularis) which he
considered to have been decorative beads. This claim was completely
ignored. In 1894, English archaeologist Worthington George Smith
discovered 200 similar perforated fossils in Bedfordshire, England, and
also speculated that their function was beads, though he made no
reference to Boucher de Perthes' find, possibly because he was unaware
of it.
In
2005, Robert Bednarik reexamined the material, and concluded that -
because all the Bedfordshire P. globularis fossils are sub-spherical and
range 10–18 mm (0.39–0.71 in) in diameter, despite this species
having a highly variable shape - they were deliberately chosen. They
appear to have been bored through completely or almost completely by
some parasitic creature (i. e., through natural processes), and were
then percussed on what would have been the more closed-off end to fully
open the hole. He also found wear facets which he speculated were
begotten from clacking against other beads when they were strung
together and worn as a necklace. In 2009, Solange Rigaud, Francisco
d'Errico and colleagues noticed that the modified areas are lighter in
colour than the unmodifed, suggesting they were inflicted much more
recently such as during excavation. They were also unconvinced that the
fossils could be confidently associated with the Acheulian artefacts
from the sites, and suggested that - as an alternative to archaic human
activity - apparent size-selection could have been caused by either
natural geological processes or 19th-century collectors favouring this
specific form.
Early modern humans and late Neanderthals (the latter especially after 60,000 years ago) made wide use of red ochre for presumably symbolic purposes as it produces a blood-like colour, though ochre can also have a functional medicinal application. Beyond these two species, ochre usage is recorded at
Olduvai
Gorge, Tanzania, where two red ochre lumps have been found; Ambrona where an ochre slab was trimmed down into a specific shape; and Terra Amata where 75 ochre pieces were heated to achieve a wide colour range from yellow to red-brown to red. These may exemplify early and isolated instances of colour preference and colour categorisation, and such practices may not have been normalised yet.
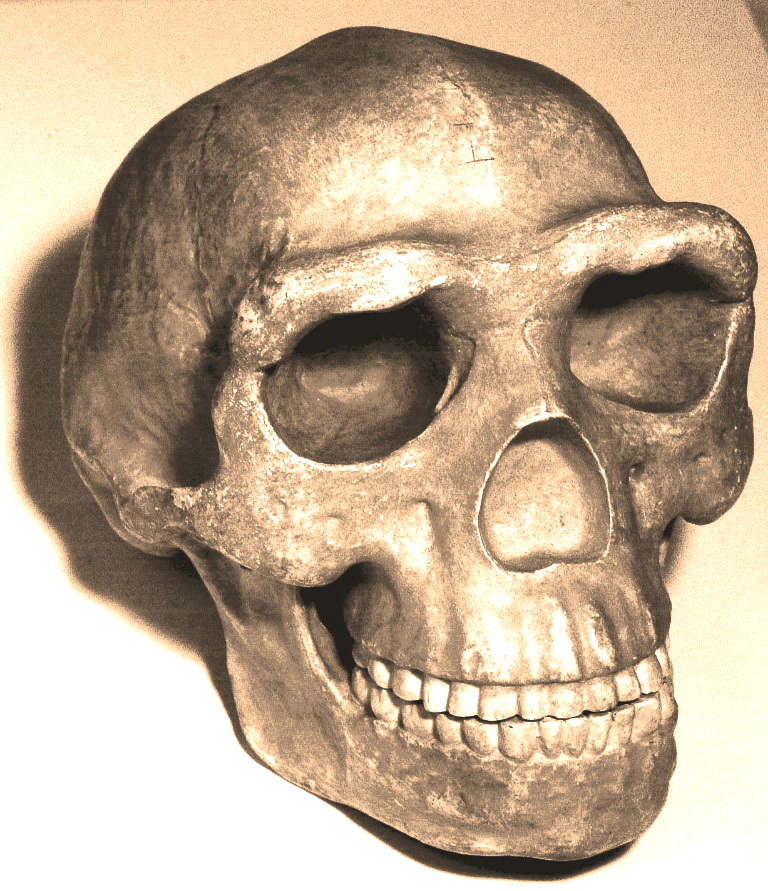
Skull
of Peking Man
TECHNOLOGY: TOOLS, FIRE, SPEARS
STONE TOOLS
The Lower Palaeolithic (Early Stone Age) comprises the Oldowan which was replaced by the
Acheulian, which is characterised by the production of mostly symmetrical hand axes. The Acheulian has a timespan of about a million years, and such technological stagnation has typically been ascribed to comparatively limited cognitive abilities which significantly reduced innovative capacity, such as a deficit in cognitive fluidity, working memory, or a social system compatible with apprenticeship. Nonetheless, the Acheulian does seem to subtly change over time, and is typically split up into Early Acheulian and Late
Acheulian, the latter becoming especially popular after 600 to 500 thousand years ago. Late Acheulian technology never crossed over east of the Movius Line into East Asia, which is generally believed to be due to either some major deficit in cultural transmission (namely smaller population size in the East) or simply preservation bias as far fewer stone tool assemblages are found east of the line.
The transition is indicated by the production of smaller, thinner, and more symmetrical hand axes (though thicker, less refined ones were still produced). At the 500,000-year-old Boxgrove site in
England - an exceptionally well-preserved site with abundance of tool
remains - thinning may have been produced by striking the hand axe near-perpendicularly with a soft hammer, possible with the invention of prepared platforms for tool making. The Boxgrove knappers also left behind large lithic flakes leftover from making hand axes, possibly with the intention of recycling them into other tools later. Late Acheulian sites elsewhere pre-prepared lithic cores ("Large Flake Blanks,"
LFB) in a variety of ways before shaping them into tools, making prepared platforms unnecessary. LFB Acheulian spreads out of Africa into West and South Asia before a million years ago and is present in Southern Europe after 600,000 years ago, but northern Europe (and the Levant after 700,000 years ago) made use of soft hammers as they mainly made use of small, thick flint nodules. The first prepared platforms in Africa come from the 450,000-year-old Fauresmith industry, transitional between the Early Stone Age
(Acheulian) and the Middle Stone Age.
With either method, knappers (tool makers) would have had to have produced some item indirectly related to creating the desired product (hierarchical organisation), which could represent a major cognitive development. Experiments with modern humans have shown that platform preparation cannot be learned through purely observational learning, unlike earlier techniques, and could be indicative of well developed teaching methods as well as self-regulated learning. At
Boxgrove, the knappers used not only stone but also bone and antler to make hammers, and the use of such a wide range of raw materials could speak to advanced planning capabilities as stoneworking requires a much different skillset to work and gather materials for than
boneworking.
The Kapthurin Formation, Kenya, has yielded the oldest evidence of blade and bladelet technology, dating to 545 to 509 thousand years ago. This technology is rare even in the Middle
Palaeolithic, and is typically associated with Upper Palaeolithic modern humans. It is unclear if this is part of a long blade-making tradition, or if blade technology was lost and reinvented several times by multiple different human species.
FIRE & CONSTRUCTION
Despite apparent pushes into colder climates, evidence of fire is scarce in the archaeological record until 400 to 300 thousand years ago. Though it is possible fire remnants simply degraded, long and overall undisturbed occupation sequences such as at Arago or Gran Dolina conspicuously lack convincing evidence of fire usage. This pattern could possibly indicate the invention of ignition technology or improved fire maintenance techniques at this time, and that fire was not an integral part of people's lives before then in Europe. In Africa, on the other hand, humans may have been able to frequently scavenge fire as early as 1.6 million years ago from natural wildfires, which occur much more often in Africa, thus possibly (more or less) regularly using fire. The oldest established continuous fire site beyond Africa is the 780,000-year-old Gesher Benot
Ya'aqov, Israel.
In Europe, evidence of constructed dwelling structures - classified as firm surface huts with solid foundations built in areas mostly sheltered from the
weather - has been recorded since the Cromerian Interglacial, the earliest example a 700,000-year-old stone foundation from Přezletice, Czech Republic. This dwelling probably featured a vaulted roof made of thick branches or thin poles, supported by a foundation of big rocks and earth. Other such dwellings have been postulated to have existed during or following the Holstein Interglacial (which began 424,000 years ago) in
Bilzingsleben, Germany; Terra Amata, France; and Fermanville and Saint-Germain-des-Vaux in Normandy. These were probably occupied during the winter, and, averaging only 3.5 m × 3 m (11.5 ft × 9.8 ft) in area, they were probably only used for sleeping in, while other activities (including
firekeeping) seem to have been done outside. Less-permanent tent technology may have been present in Europe in the Lower Paleolithic.
SPEARS
The appearance of repeated fire usage - earliest in Europe from Beeches Pit, England, and Schöningen,
Germany - roughly coincides with hafting technology (attaching stone points to spears) best exemplified by the Schöningen spears. These nine wooden spears and spear
fragments - in addition to a lance, and a double-pointed stick - date to 300,000 years ago and were preserved along a lakeside. The spears vary from 2.9–4.7 cm (1.1–1.9 in) in diameter, and may have been 210–240 cm (7–8 ft) long, overall similar to present day competitive javelins. The spears were made of soft spruce wood, except for spear 4 which was (also soft) pine wood. This contrasts with the Clacton spearhead from
Clacton-on-Sea, England, perhaps roughly 100,000 years older, which was made of hard yew wood. The Schöningen spears may have had a range of up to 35 m (115 ft), though would have been more effective short range within about 5 m (16 ft), making them effective distance weapons either against prey or predators. Besides these two localities, the only other site which provides solid evidence of European spear technology is the 120,000-year-old Lehringen site, Germany, where a 238 cm (8 ft) yew spear was apparently lodged in an elephant. In Africa, 500,000-year-old points from Kathu Pan 1, South Africa, may have been hafted onto spears. Judging by indirect evidence, a horse scapula from the 500,000-year-old Boxgrove shows a puncture wound consistent with a spear wound. Evidence of hafting (in both Europe and Africa) becomes much more common after 300,000 years.
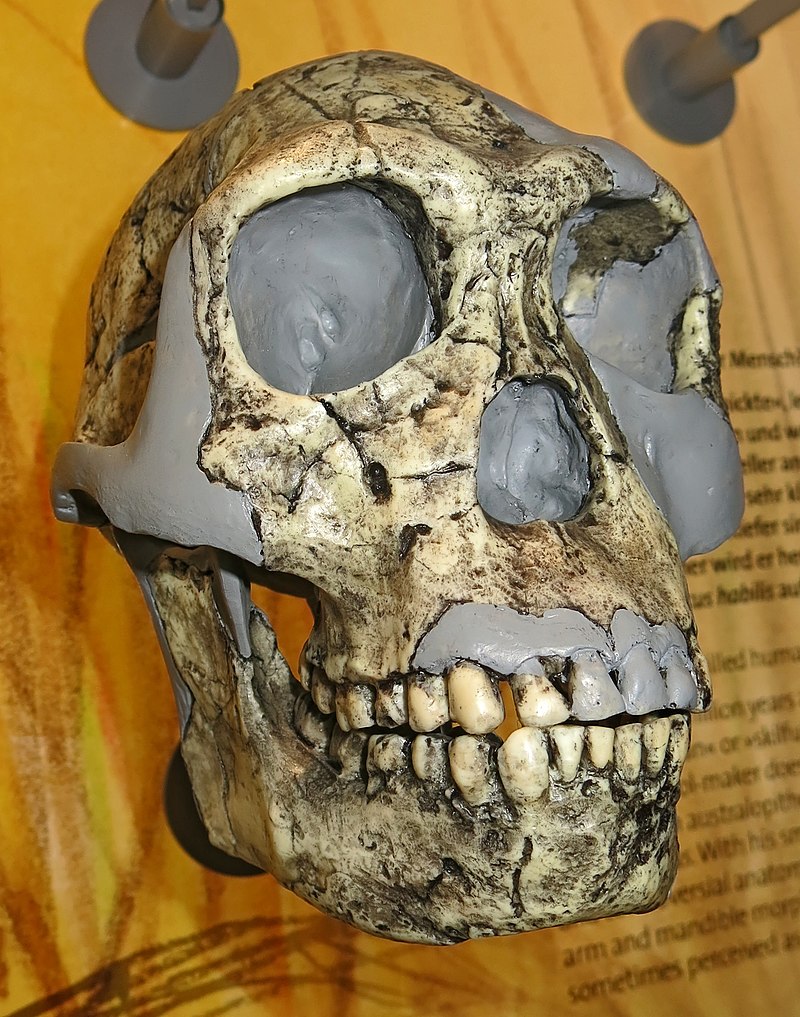
Reconstructed
skull of Homo Neanderthalis, brows less pronounced.
KANIS
REX - HOMO SAPIENS SUPERIOR
Man
took millions of years to develop from the apes, into
Homo Sapiens Sapiens, king of the primates until John
Storm was accidentally injected with a CRISPR
virus developed by Brazilian scientists working for the secret society; NeuWelt
Rittertum. The amateur anthropologist was later forced to
undo damaging alterations to his DNA, in so doing creating a new
species, physically and mentally superior to Homo S. Sapiens; named Homo
Sapiens Superior, or Kanis
Rex. These modifications were sufficient in scope to place him well
and truly in a new class. Progressing from Homo Sapiens Sapiens, to
become the king of men genetically.
The
modifications, only possible with the CyberCore Genetica™ super
computer, combined with the BioCore™
brain implant - and Hal's
AI,
were not just to his mental and muscular capabilities, but also to his
physical frame; his skeleton. Which is now stronger and lighter. All of
which in the natural world, would have taken a million or so years to
have achieved, assuming there would have been a clear natural selection advantage.

Sahelanthropus
Ardipithecus
Australopithecus
A. anamensis
A. afarensis
A. bahrelghazali
A. africanus
A. garhi
A. sediba
Kenyanthropus
Paranthropus robustus
Homo habilis
H. floresiensis
H. erectus
H. e. georgicus
H. cepranensis
H. antecessor
H. heidelbergensis
H. naledi
H. helmei
Homo neanderthalensis
Homo sapiens
H. s. idaltu
Homo Sapiens Sapiens
Homo
Sapiens Superior (Kanis
Rex)

EVOLUTION
Humans are apes (superfamily Hominoidea). The lineage of apes that eventually gave rise to humans first split from gibbons (family Hylobatidae) and orangutans (genus Pongo), then gorillas (genus
Gorilla), and finally, chimpanzees and bonobos (genus Pan). The last split, between the human and
chimpanzee - bonobo lineages, took place around 8 - 4 million years ago, in the late Miocene epoch. During this split, chromosome 2 was formed from the joining of two other chromosomes, leaving humans with only 23 pairs of chromosomes, compared to 24 for the other apes. Following their split with chimpanzees and bonobos, the hominins diversified into many species and at least two distinct genera. All but one of these
lineages - representing the genus Homo and its sole extant species Homo
sapiens - are now extinct.
The genus Homo evolved from Australopithecus. Though fossils from the transition are scarce, the earliest members of Homo share several key traits with Australopithecus. The earliest record of Homo is the 2.8 million-year-old specimen LD 350-1 from Ethiopia, and the earliest named species are Homo habilis and Homo rudolfensis which evolved by 2.3 million years ago. H. erectus (the African variant is sometimes called H. ergaster) evolved 2 million years ago and was the first archaic human species to leave Africa and disperse across Eurasia. H. erectus also was the first to evolve a characteristically human body plan. Homo sapiens emerged in Africa around 300,000 years ago from a species commonly designated as either H. heidelbergensis or H. rhodesiensis, the descendants of H. erectus that remained in Africa. H. sapiens migrated out of the continent, gradually replacing or interbreeding with local populations of archaic humans.
Humans began exhibiting behavioral modernity about 160,000-70,000 years ago, and possibly earlier.
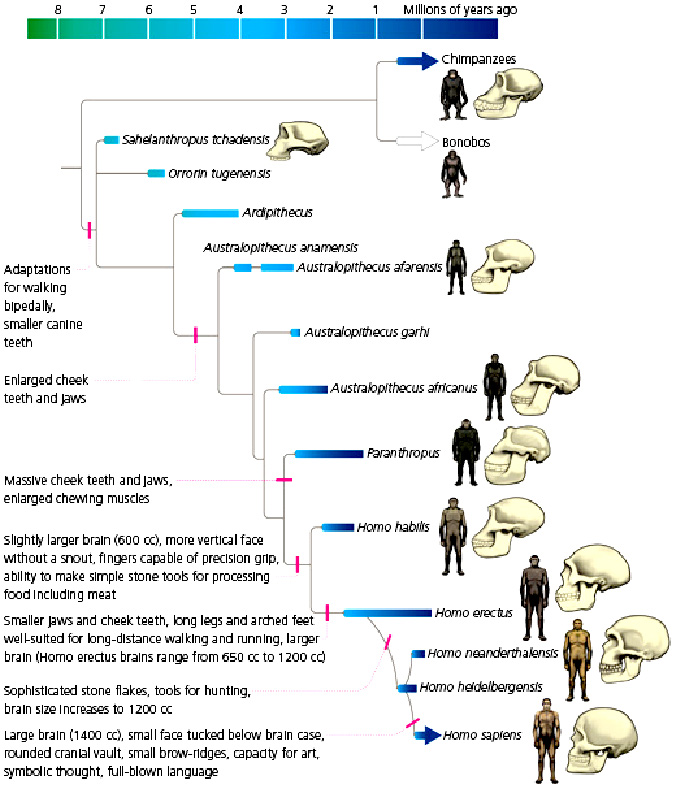
The "out of Africa" migration took place in at least two waves, the first around 130,000 to 100,000 years ago, the second (Southern Dispersal) around 70,000 to 50,000 years ago. H. sapiens proceeded to colonize all the continents and larger islands, arriving in Eurasia 125,000 years ago, Australia around 65,000 years ago, the Americas around 15,000 years ago, and remote islands such as Hawaii, Easter Island, Madagascar, and New Zealand between the years 300 and 1280 CE.
Human evolution was not a simple linear or branched progression but involved interbreeding between related species.
Genomic research has shown that hybridization between substantially diverged lineages was common in human evolution.
DNA evidence suggests that several genes of Neanderthal origin are present among all non sub-Saharan African populations, and Neanderthals and other hominins, such as Denisovans, may have contributed up to 6% of their genome to present-day non sub-Saharan African humans.
Human evolution is characterized by a number of morphological, developmental, physiological, and behavioral changes that have taken place since the split between the last common ancestor of humans and
chimpanzees. The most significant of these adaptations are obligate bipedalism, increased brain size and decreased sexual dimorphism (neoteny). The relationship between all these changes is the subject of ongoing debate.

The
Cup of Christ is the Holy Grail, that has never been found, in all
searches through the ages.